Анализ белка (в том числе моноклонального антитела для медицинского применения по ОФС 1.7.1.0014) в биофармацевтике в первую очередь направлен на подтверждение соответствия структуры полученного продукта структуре ожидаемой. Основные потенциальные проблемы возникают, как правило, на уровне первичной структуры белка; в зависимости от их источника (генетического, трансляционного или пост-трансляционного) возможны нарушения различного рода – измененные участки аминокислотной последовательности, частичные аминокислотные замены, модифицированные отдельные остатки. Для выявления этих проблем может быть использован широкий набор аналитических подходов – от анализа нативного белка до анализа его коротких фрагментов, содержащих проблематичные участки. Анализ фрагментов с применением масс-спектрометрии высокого разрешения в конечном счете является решающим, так как при этом не только констатируется факт несоответствия первичной структуры, но и проводится идентификация изменения, включая локализацию модифицированного остатка.
Расщепление белка до пептидных фрагментов, как правило, удобнее проводить ферментативно, так как при химической фрагментации возможно образование дополнительно модифицированных побочных продуктов. Анализ образовавшихся пептидов проводится с использованием жидкостной хроматографии / масс-спектрометрии. Первичная задача здесь – получение репрезентативного списка найденных (и идентифицированных) протеолитических продуктов. Мы никогда не полагаемся на полностью автоматизированную обработку данных, так как при использовании программной процедуры результаты идентификации будут ограничены заранее задаваемым списком возможных изменений. Для гарантированного выявления всех значимых компонентов «ручная» обработка обязательна.
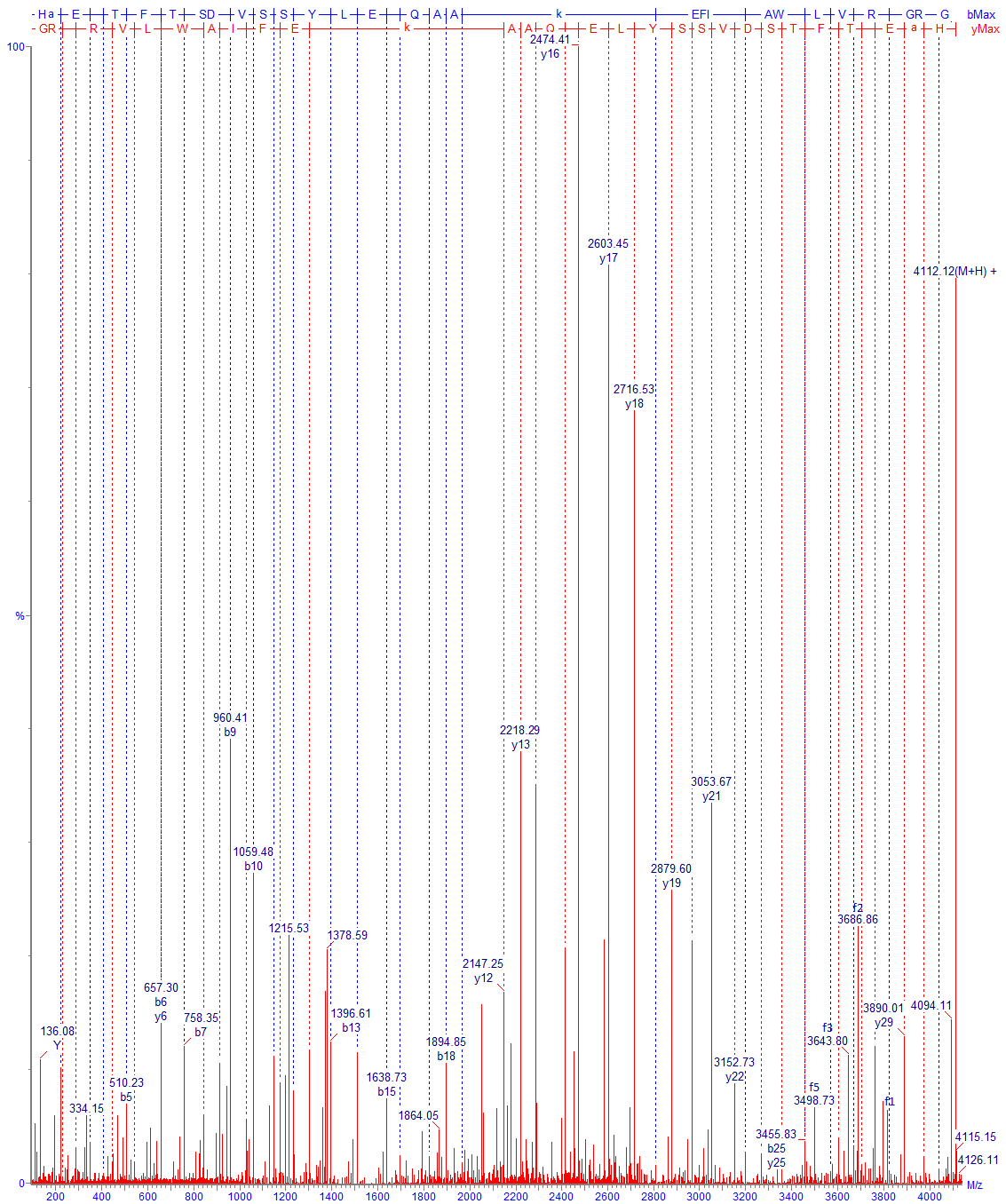
В состав основной части работы фактически входит «пептидное картирование» (ОФС.1.7.2.0035.18) исследуемого белка (Рис. 1, Рис. 2). Исторически метод пептидного картирования включал фрагментацию белка, разделение полученных фрагментов хроматографическими либо электрофоретическими методами и, наконец, качественное (визуальное) сравнение расположения и высоты пиков среди наборов, полученных для анализируемого и стандартного образцов. Использование методов масс-спектрометрии кардинально поменяло суть метода, добавив смысловую нагрузку: ранее метод оперировал на уровне параметров абстрактных пиков, сейчас – на уровне структуры идентифицируемых компонентов. Кроме того, совершенно необходимое ранее условие о наличии стандартного образца сейчас является необязательным (но часто желательным).
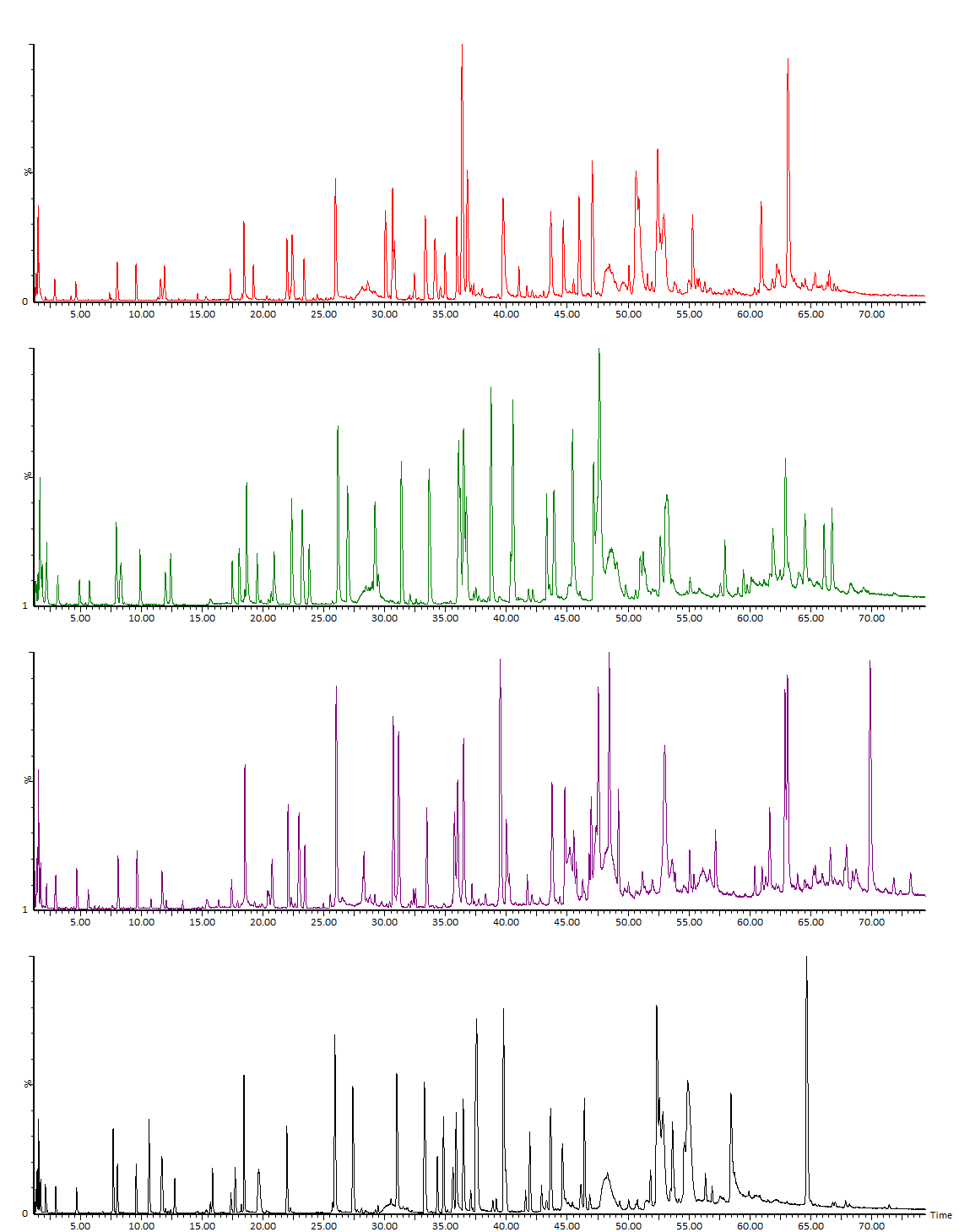
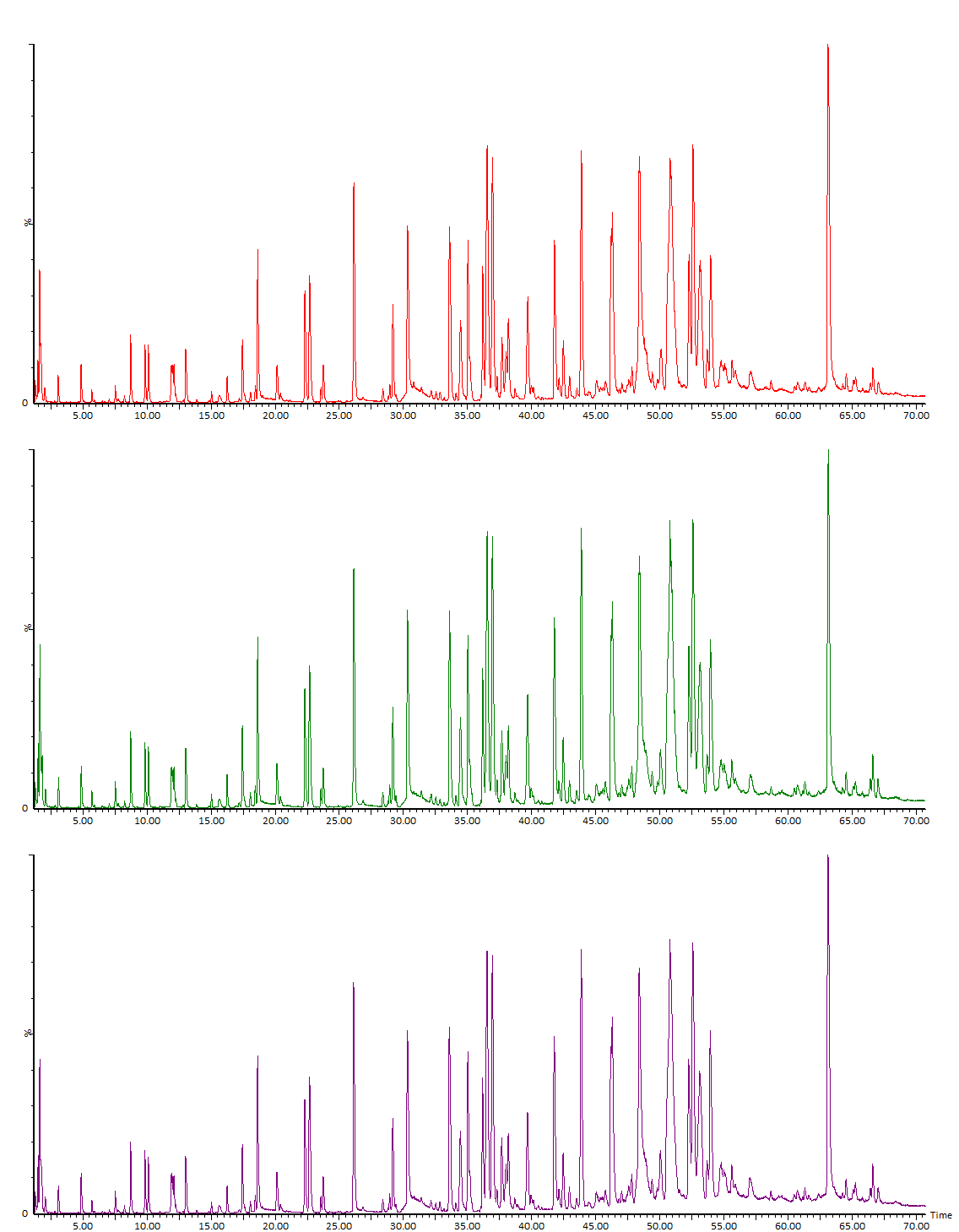
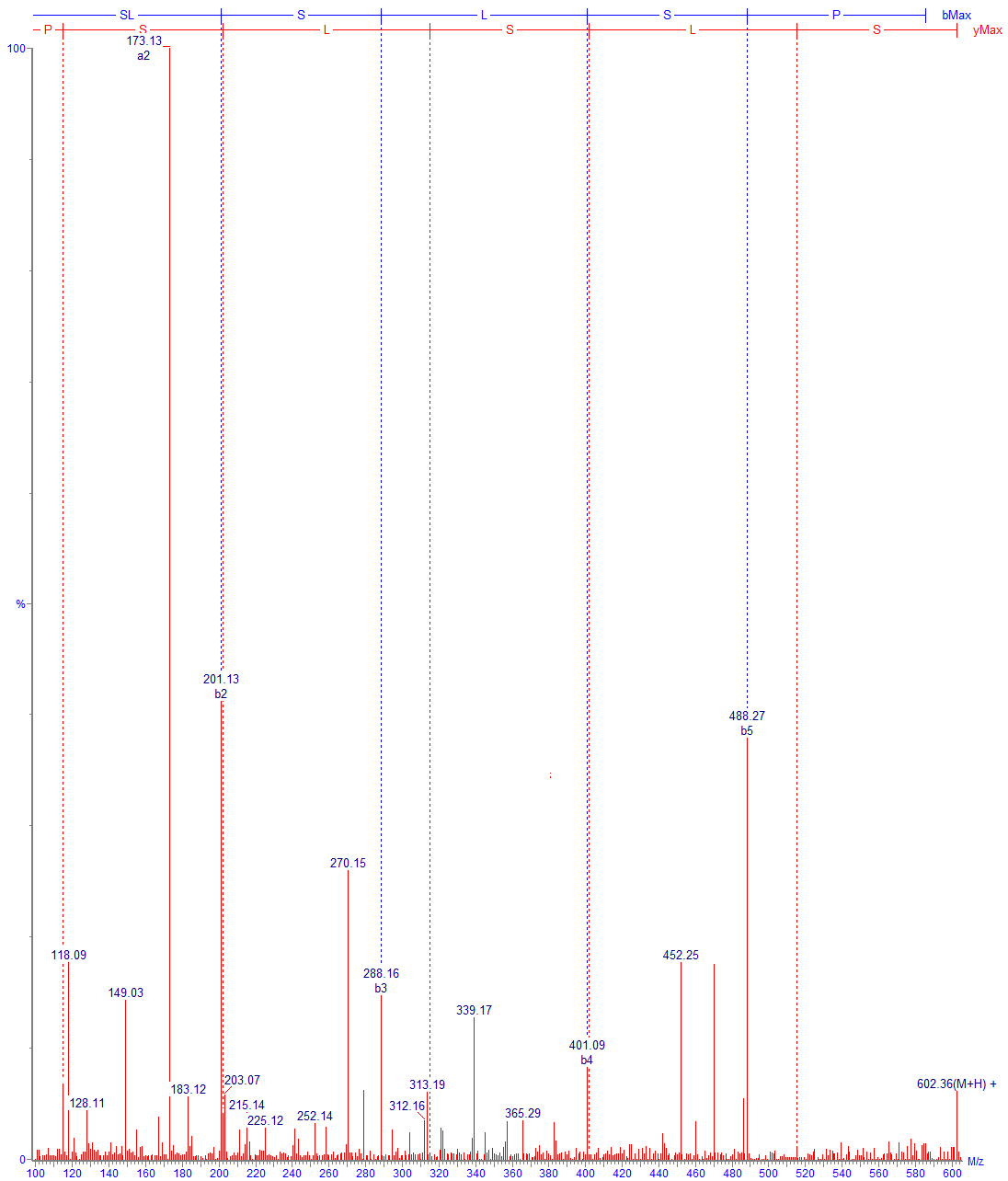
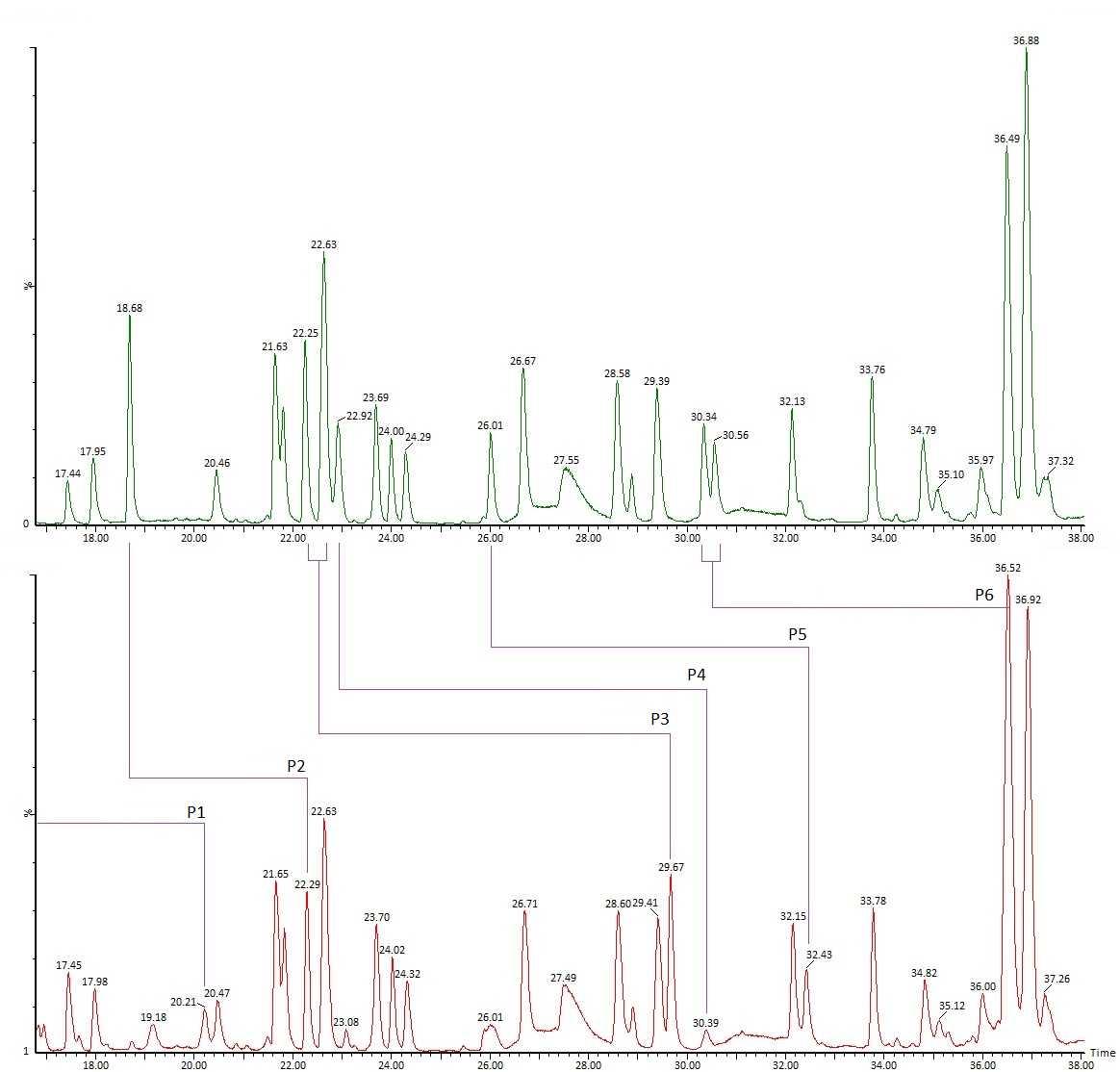
Идентификация пептидов проводится по значениям точных масс и по спектрам фрагментации. При анализе типичных моноклональных антител обычно наш список идентифицированных пептидов содержит порядка полутора сотен компонентов. В него включаются продукты полного гидролиза, частичного и неспецифического, кроме того, частично – основные гликопептиды и продукты модификации, дополнительные данные о модификациях мы указываем в отдельных разделах Отчёта. На практике после тщательного анализа в большинстве случаев видны все ожидаемые участки полной цепи белка; формально, однако, перекрывание последовательности будет немного ниже 100 % (обычно 95-99 %), так как метод не позволяет зарегистрировать короткие гидрофильные пептиды, не связывающиеся с обращенно-фазовым сорбентом. В таких случаях степень перекрывания иногда можно увеличить, используя более мягкие условия гидролиза. В редких случаях определенный пептид может не детектироваться ввиду крайне низкой ионизируемости, как, например, отдельные гликопептиды с очень высоким содержанием сиаловой кислоты. Проблему можно решать специфическими подходами, в примере выше можно использовать предварительную обработку белка сиалидазой (альфа-2-3,6,8,9 нейраминидазой А) для снятия связанных остатков сиаловой кислоты.
Проверка последовательности цепи белка автоматически включает анализ N-/C- концевых участков. В случае моноклональных антител на этих участках обычно наблюдается несколько характерных изменений. C-концевой лизин тяжелой цепи, как правило, частично отсутствует, кроме того, могут наблюдаться варианты, где отсутствует и вторая концевая аминокислота, а предшествующая ей (обычно пролин) находится в форме амида (Рис. 3). В тех случаях, если N-концевой остаток цепи – глутамин или глутаминовая кислота, обычно происходит циклизация с образованием пироглутамата (Рис. 4). Иногда регистрируются продукты с частично неотщепившимся участком сигнального пептида (Рис. 5).
Характеристика гликоформ белка – отдельная задача, на которой мы подробнее остановимся в соответствующем разделе.
К наиболее распространенным нежелательным пост-трансляционным модификациям относятся окисление метионина и дезамидирование аспарагина. Скорость, с которой модифицируются отдельные остатки, может различаться на порядки: она определяется различными факторами, в том числе расположением модифицируемого остатка в молекуле белка и его пространственной доступностью, а также окружением. Так, в цепях IgG имеется несколько участков, особенно чувствительных к окислению или дезамидированию.
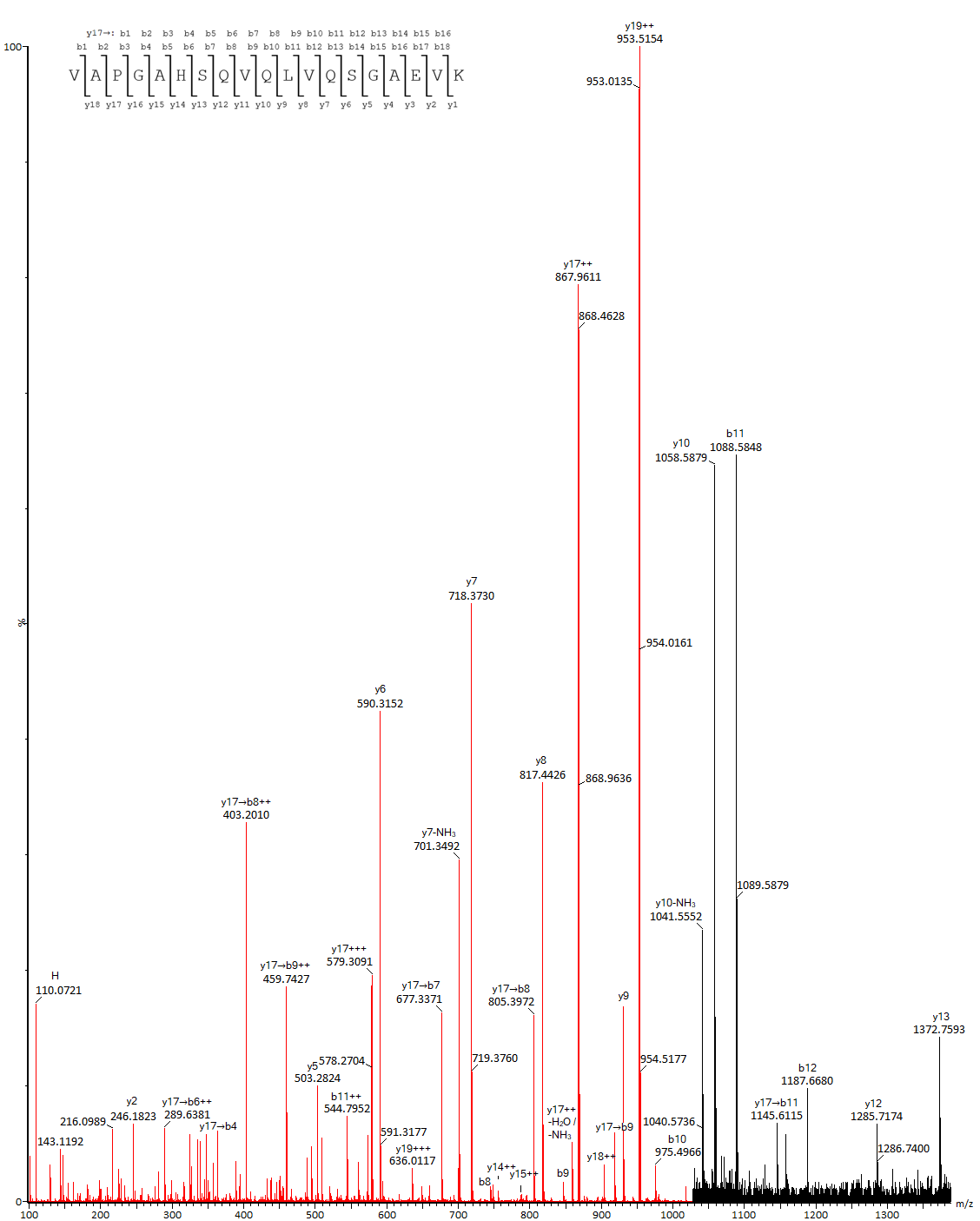
Типичным маркером окисления является пептид DTLMISR, образующийся при гидролизе IgG трипсином (Рис. 6). С меньшей скоростью окисляется метионин на другом консервативном участке, SVMHEAL. Состояние метионина в вариабельных областях может быть различным в зависимости от окружения и положения в цепи.
Подверженность остатков аспарагина дезамидированию дополнительно определяется особыми факторами. Дезамидирование может проходить по двум основным механизмам; в нормальных условиях преобладает путь с образованием аспартил-сукцинимида и последующим его гидролизом до аспартата и изо-аспартата. Для скорого прохождения этой реакции необходимо, чтобы в цепи за аспарагином следовал глицин, поэтому аспарагины на участках …NG… весьма подвержены модификации. В консервативных областях IgG есть два-три таких участка, из которых в первую очередь выделяется GFYPSDIAVEWESNGQPENNYK, получивший неформальное название PENNY-пептид (Рис. 7). В этом пептиде три остатка аспарагина и один глутамин. Первый аспарагин (ESNGQ) практически всегда частично дезамидирован (обычно в пределах 1 – 10 %), причем обычно детектируется весь набор возможных продуктов: сукцинимид, изо-аспартат и аспартат. Также часто наблюдается модификация и второго остатка аспарагина (PENNY); в этом случае механизм дезамидирования иной и среди продуктов детектируется только аспартат. Другой участок …NG… включается в триптический пептид VVSVLTVLHQDWLNGK, где аспарагин также легко подвержен модификации, причем обычно преобладающим продуктом оказывается аспартил-сукцинимид. По результатам анализа различных моноклональных антител, степень дезамидирования других остатков аспарагина / глутамина в большинстве случаев оказывается меньшей, однако возможны исключения, когда за счет особого окружения скорость модификации определенной аминокислоты существенно возрастает. В качестве примера можно привести трастузумаб, где Asn30L на участке ASQDVNTAVAWYQQKPGK часто дезамидирован в большей степени, чем Asn387H PENNY-пептида.

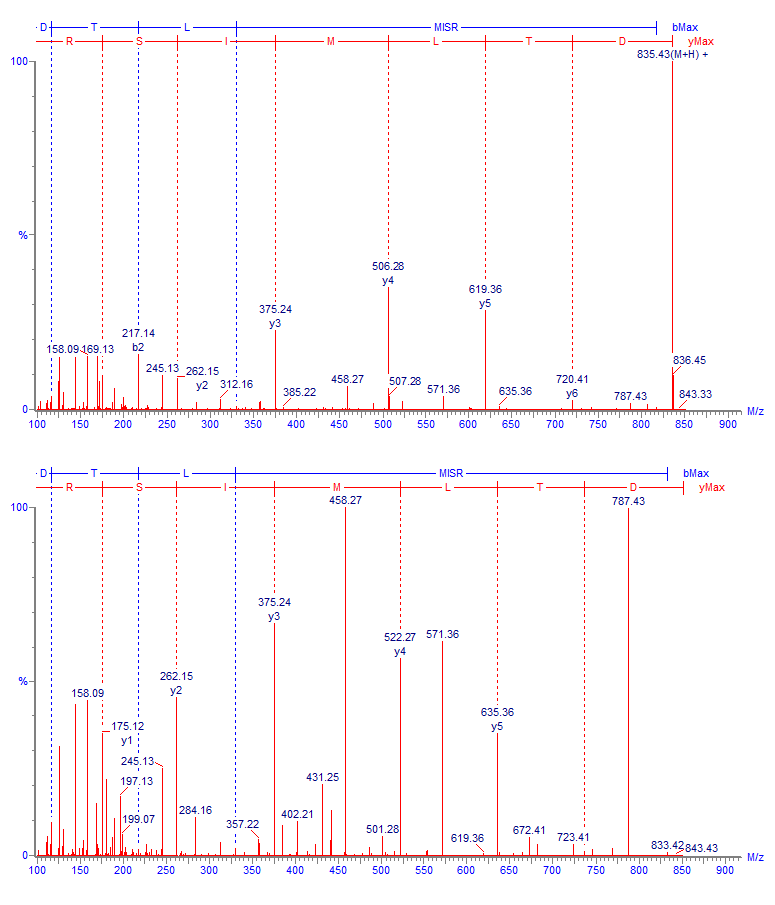
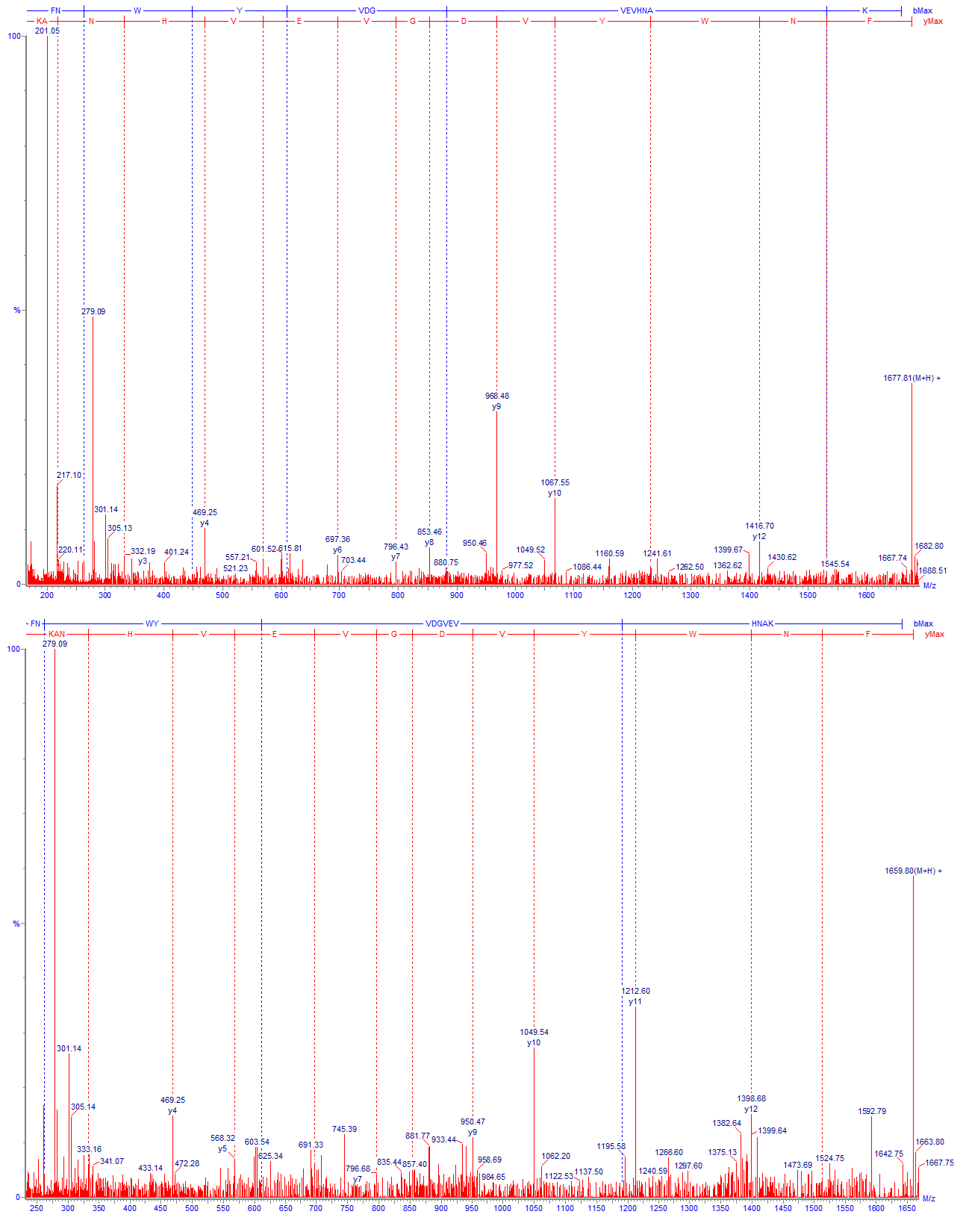
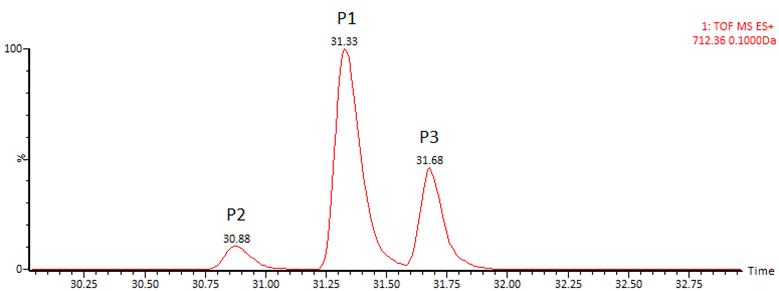
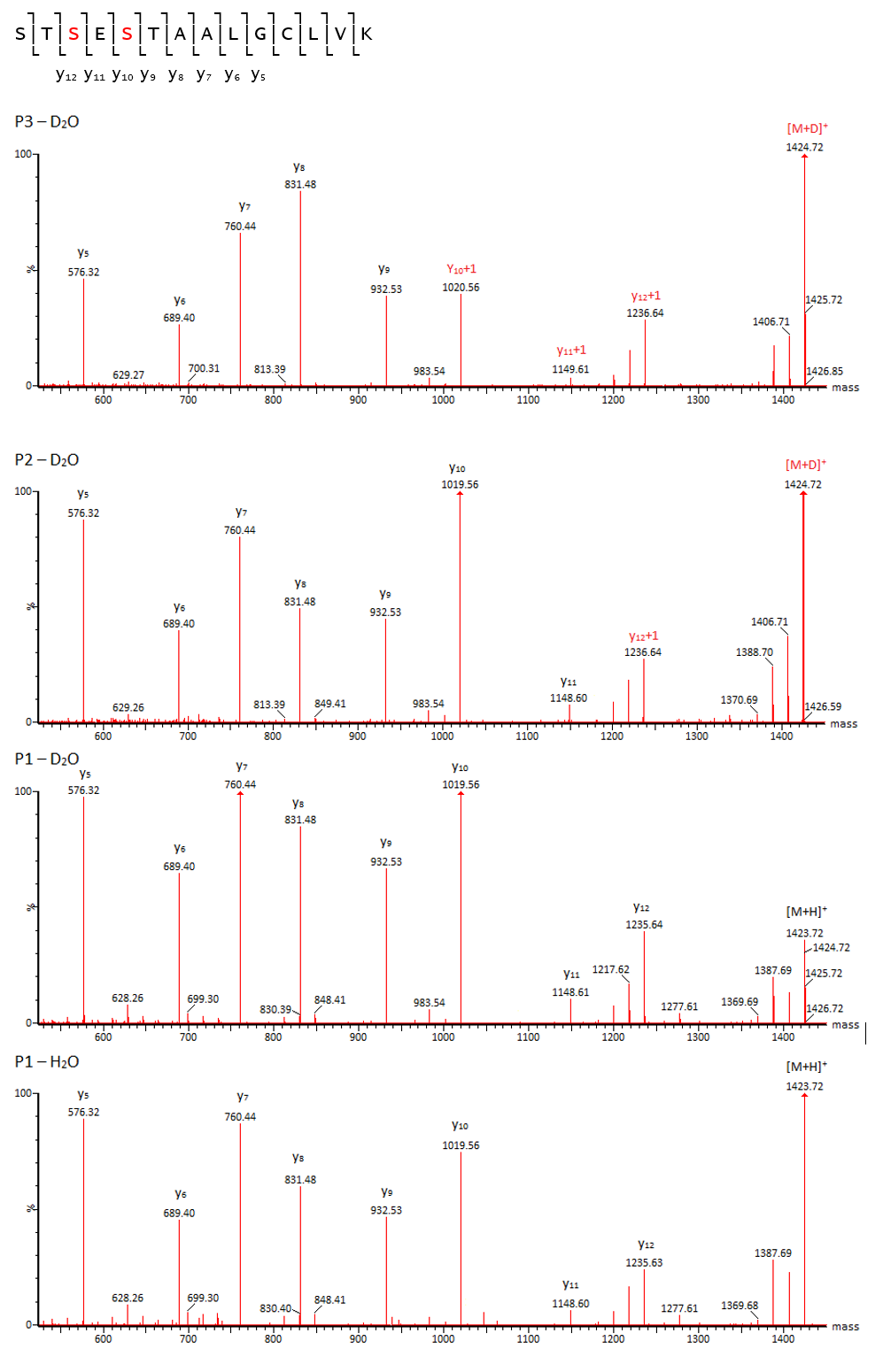
Изо-аспартат в цепи белка также может образовываться из остатков аспарагиновой кислоты. Механизм аналогичен пути дезамидирования аспарагина с образованием isoAsp, однако в данном случае реакция, как правило, протекает намного медленнее и содержание образовавшегося продукта обычно находится на очень низком уровне. В качестве примера (Рис. 8) можно привести образование изо-аспартата на консервативном участке IgG FNWYVDGVEVHNAK: оно наблюдается практически всегда во всех препаратах, но в весьма слабой степени (0.1 – 1 %). Кроме формы с изо-аспартатом здесь также детектируется промежуточный продукт с аспартил-сукцинимидом, что подтверждает идентификацию (Рис. 9).
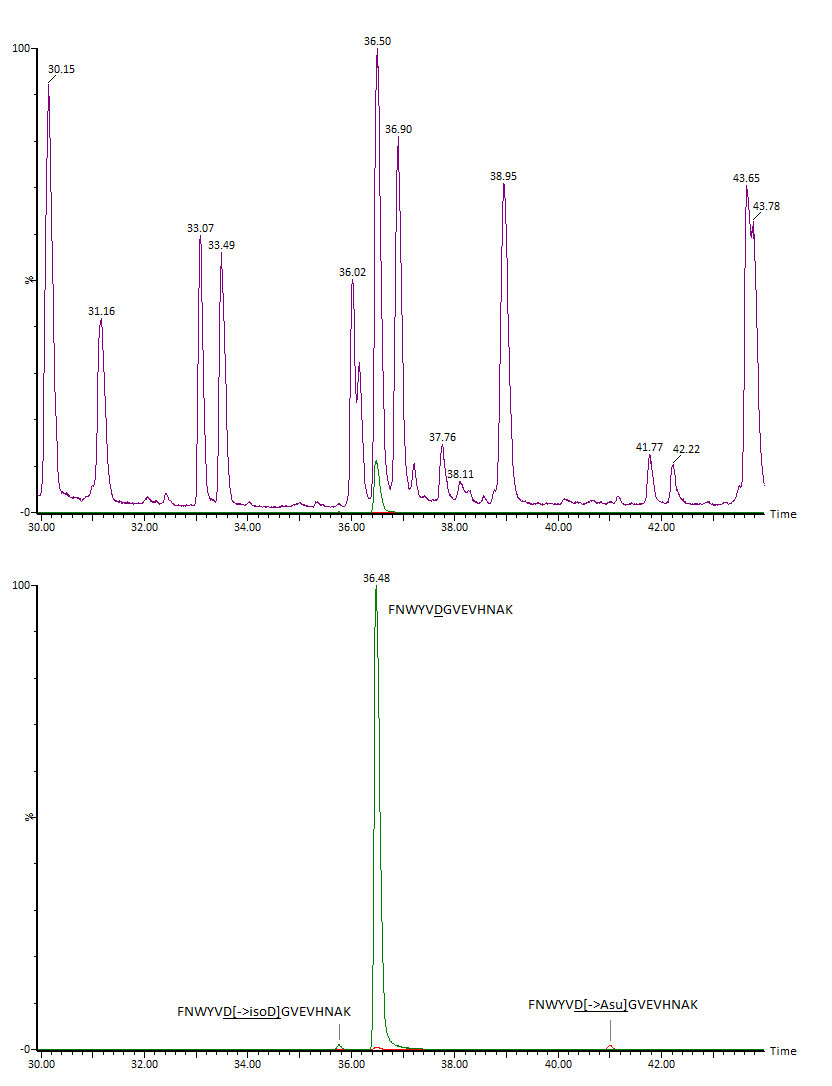
Верхняя хроматограмма – MS(ESI+)TIC, нижняя – по выделенным ионам m/z 559.94 (D‑> isoD) и m/z 553.93 (D‑> Asu).
Другая существенная пост-трансляционная модификация – рацемизация. В норме она обычно не проявляется, однако при нарушении условий производства или хранения содержание модифицированных форм может быть значительным. Рацемизации в цепи белка наиболее подвержены остатки серина и аспартата; реакция относительно легко проходит в слабощелочной среде и может быть выявлена в экспериментах по ускоренной деградации. При рацемизации пептидов образуются диастереомеры, которые могут быть разделены хроматографически на нехиральных сорбентах. В обычных условиях переход аминокислоты из L- в D-форму не приводит к изменению массы, однако при проведении реакции в среде D2O при рацемизации происходит включение дейтерия, что позволяет средствами масс-спектрометрии установить как сам факт модификации, так и положение модифицируемого остатка (Рис. 10,11).
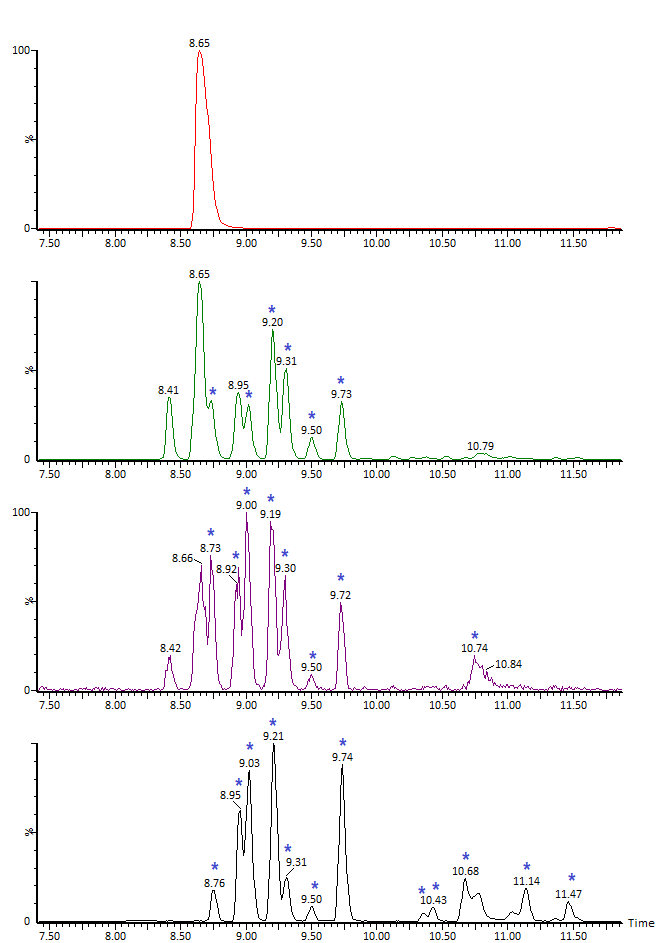
Эксперименты по ускоренной деградации белка относятся к исследованиям его стрессовой стабильности – необходимой стадии разработки биофармацевтических препаратов. Здесь возможна проверка многочисленных параметров, из которых обычно в первую очередь существенны устойчивость к окислению и устойчивость к изменению pH (Рис. 12, Рис. 13). В зависимости от условий изменения в структуре белка могут быть весьма масштабными; в частности, щелочная обработка приводит к деградации дисульфидов и O-гликанов, в том числе сопровождающейся разрывами полипептидной цепи, дезамидированию остатков Asn/Gln, рацемизации Ser/Asp и другим.
В том случае, если анализируемый белок содержит дисульфидные связи, встаёт задача определения как их положения, так и состояния (степень замкнутости, наличие продуктов рекомбинации и др.). В общем виде задача решается через анализ пептидов, полученных после гидролиза белка в невосстанавливающих условиях. Для точной локализации всех остатков Cуs, образующих дисульфиды, обычно необходимо, чтобы анализируемые пептиды содержали не более одного цистина на молекулу. В ряде случаев этого можно добиться путем выбора подходящего протеолитического фермента, иначе возможно использование более сложных подходов, таких как секвенирование с использованием экзопептидаз или анализ частично восстановленного белка.
Часто объектами исследований являются небольшие терапевтически значимые белки и тяжелые пептиды, такие как инсулины или синтетические белки, построенные на основе GLP (Glucagon-like peptide, Глюкагоноподобный пептид)
Кликните, чтобы показать таблицы с наименованиями инсулинов и глутидов
| INN | МНН | Brand Name | Наименование препарата |
| Semaglutide | Семаглутид | Ozempic, Wegovy | Оземпик, Вегови, Семавик |
| Liraglutide | Лираглутид | Victoza | Виктоза |
| Dulaglutide | Дулаглутид | Trulicity | Трулисити |
| Exenatide | Эксенатид | Byetta, Bydureon | Баета |
| Lixisenatide | Ликсисенатид | Adlyxin | |
| Teduglutide | Тедуглутид | Gattex, Revestive | Гэттестив |
| Tirzepatide | Тирзепатид | Mounjaro | Мунджаро (Мунжаро) |
| INN | МНН | Brand Name | Наименование препарата |
| Insulin lispro | инсулин лизпро | Humalog | Хумалог, РинЛиз |
| Insulin aspart | инсулин аспарт | Novorapid, Fiasp | Новорапид, Фиасп, РинФаст |
| Insulin glulisine | инсулин глулизин | Apidra | Апидра, РинГлузин |
| Insulin glargine | инсулин гларгин | Lantus, Toujeo | Лантус, РинГлар, Туджео |
| Insulin detemir | инсулин детемир | Levemir | Левемир |
| Insulin degludec | инсулин деглудек | Tresiba | Тресиба |
| Insulin degludec + aspart | инсулин деглудек + инсулин аспарт | Ryzodeg | Райзодег |
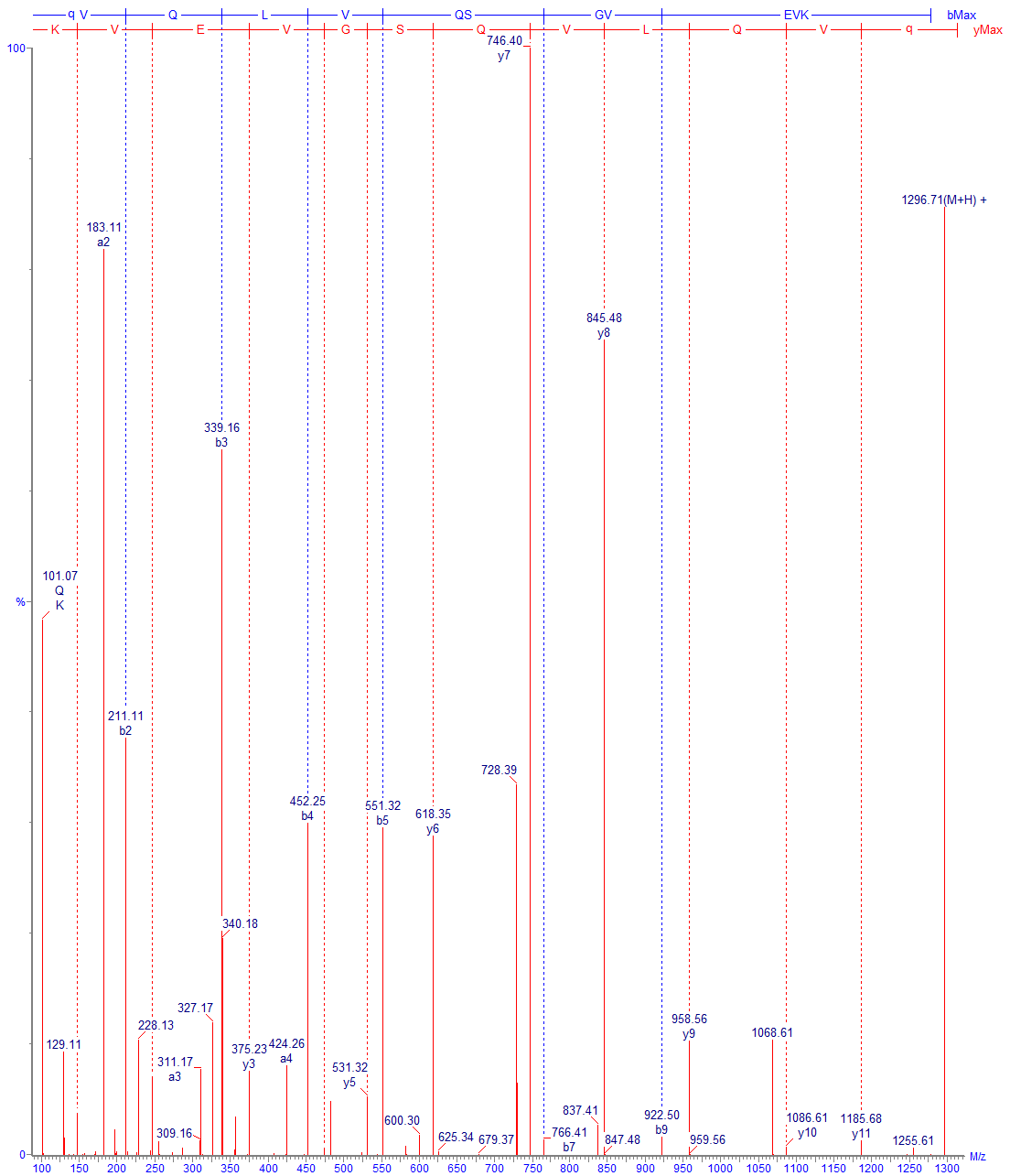
В некоторых типах инсулинов присутствуют искусственно введённые заместители. Например, в инсулине деглудек имеется остаток гексадекандиоевой кислоты, присоединенный через L-γ-глутамат. В тирзепатиде в основную цепь встроены два остатка α-аминоизомасляной кислоты, а также имеется остаток эйкозандиоевой кислоты, присоединенный через L-γ-глутамат и две молекулы [2-(2-аминоэтокси)этокси]уксусной кислоты.
Хотя в таких случаях обычно по регламенту требуется проведение пептидного картирования, подтверждение аминокислотной последовательности возможно уже по спектрам фрагментации полных цепей (Рис. 14,15,16).
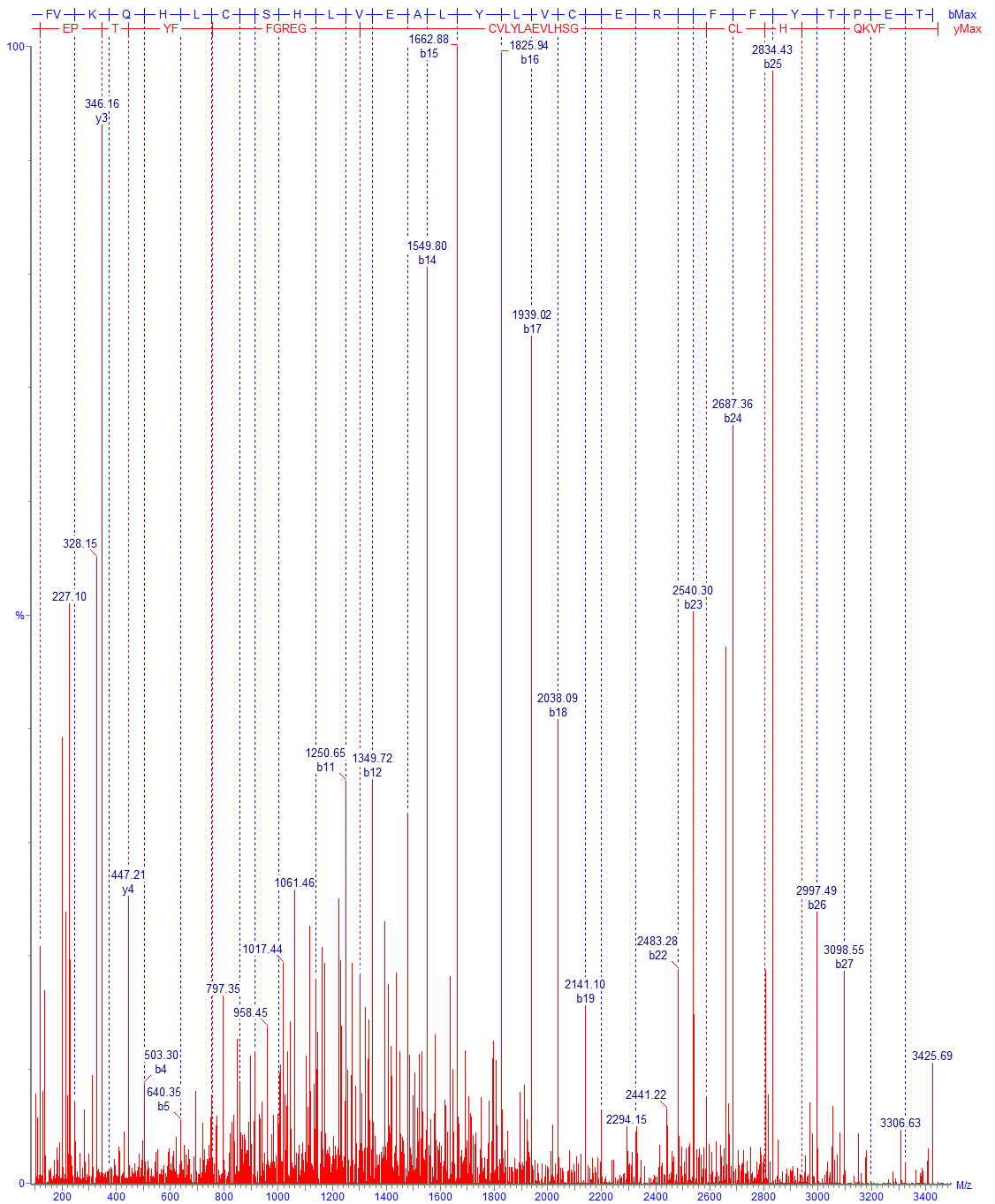
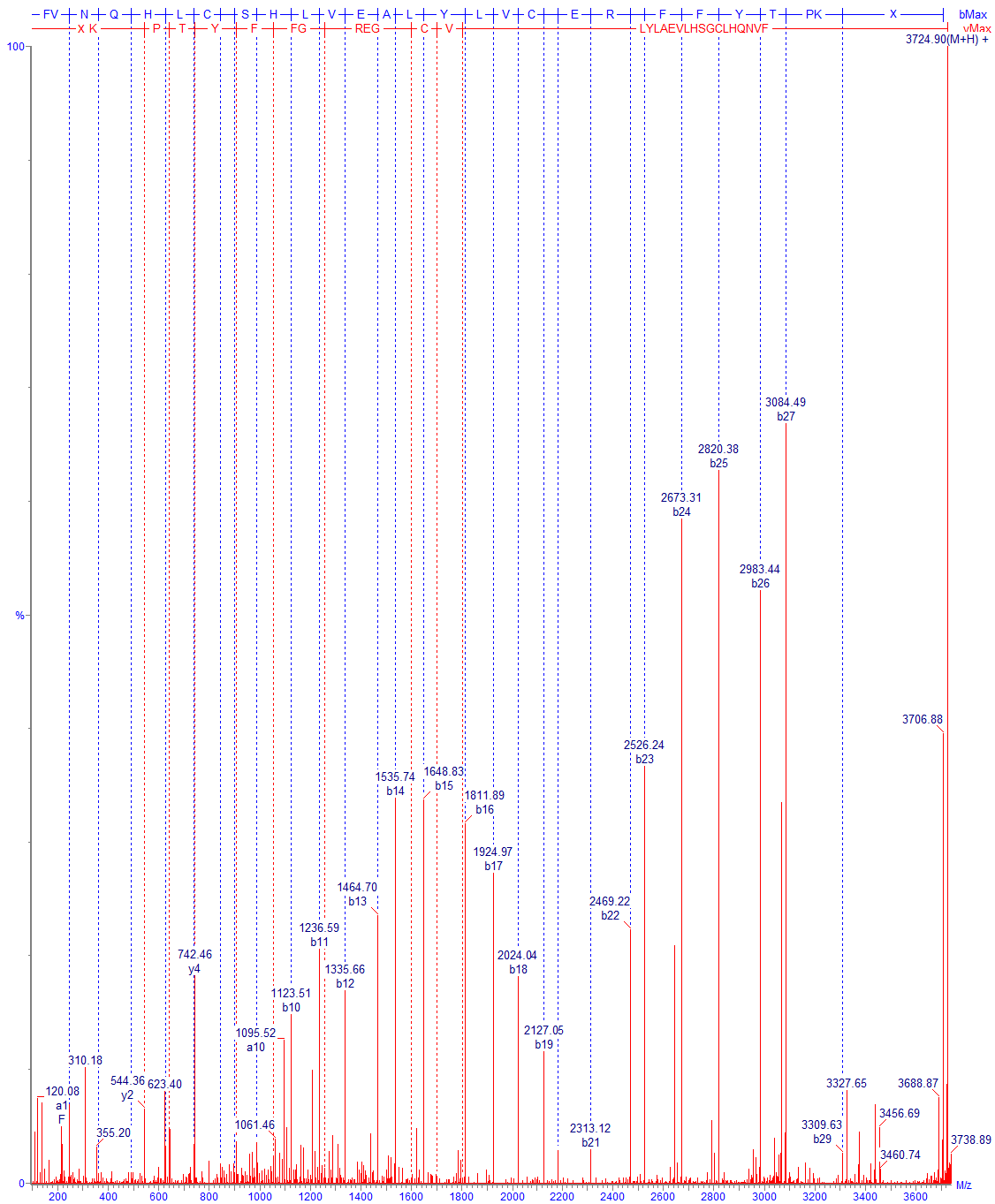
Технически успех анализа протеолитических фрагментов белка определяется рядом факторов. Это включает как выбор оптимальных условий гидролиза, так и выбор оптимальной пробоподготовки. В большинстве случаев (включая IgG) протеолиз нативного белка идет крайне медленно, поэтому предварительно белок необходимо тем или иным способом денатурировать. Предварительные стадии могут включать восстановление, а также блокировку лабильных групп (обычно цистеинов). При проведении пробоподготовки следует учитывать, что возможны потери продуктов как вследствие плохой растворимости, так и вследствие других факторов, например возможной сорбции на стенках. Условия пробоподготовки должны быть подобраны таким образом, чтобы при проведении процедуры не образовывались новые модифицированные продукты; особенно трудно полностью избежать частичное дезамидирование аспарагина на чувствительных участках, а также частичную рекомбинацию при анализе дисульфидов.
Мы используем свою базовую методику, с одной стороны, устоявшуюся и хорошо проверенную, с другой – легко адаптируемую под особенности анализа конкретного объекта. При её применении обычно мы видим все ожидаемые продукты (что говорит об отсутствии возможных потерь при пробоподготовке). Образование модифицированных форм можно оценить так: окисление метионина – практически отсутствует (<0.2%), дезамидирование аспарагина – менее 1-1.5 % (на участках GFYPSDIAVEWESNGQPENNYK и VVSVLTVLHQDWLNGK), образование смешанных дисульфидов – в пределах 2 % (обычно < 0.5 %).
В настоящее время известно более 200 лекарственных препаратов на основе моноклональных антител.
Прежде всего это препараты для лечения онкологических заболеваний (кликните, чтобы увидеть таблицу):
| INN | МНН | Brand Name | Наименование препарата |
| Adebrelimab | Адебрелимаб | Adebrelimab | Адебрелимаб |
| Ado-trastuzumab emtansine | Трастузумаб эмтанзин | Kadcyla | Кадсила |
| Alemtuzumab | Алемтузумаб | Lemtrada; MabCampath, Campath-1H | Лемтрада Мабкампат |
| Amivantamab | Амивантамаб | Rybrevant | Рибревант |
| Atezolizumab | Атезолизумаб | Tecentriq | Тецентрик |
| Avelumab | Авелумаб | Bavencio | Бавенсио |
| Belantamab mafodotin (belantamab mafodotin-blmf) | Белантамаб мафодотин | Blenrep | Бленреп |
| Bevacizumab | Бевацизумаб | Avastin | Авастин, Авегра, Бевацизумаб, Версаво, Стибевара |
| Blinatumomab | Блинатумомаб | Blincyto | Блинцито |
| Brentuximab vedotin | Брентуксимаб ведотин | Adcetris | Адцетрис |
| Cadonilimab | Кадонилимаб | 开坦尼 | Кадонилимаб |
| Camrelizumab | Камрелизумаб | AiRuiKa | Арейма |
| Catumaxomab | Катумаксомаб | Removab | Ремоваб |
| Cemiplimab (cemiplimab-rwlc) | Цемиплимаб | Libtayo | Либтайо |
| Cetuximab | Цетуксимаб | Erbitux | Эрбитукс, Цетуксимаб |
| Cetuximab saratolacan | Цетуксимаб саратолакан | Akalux® IV Infusion | |
| Cosibelimab | косибелимаб | Unloxcyt | Анлоксит |
| Daratumumab | Даратумумаб | Darzalex | Дарзалекс |
| Dinutuximab | Динутуксимаб | Unituxin | Унитуксин |
| Dinutuximab beta | Динутуксимаб бета | Qarziba | Карзиба |
| Disitamab vedotin | Диситамаб ведотин | Aidixi | |
| Dostarlimab | Достарлимаб | Jemperli | Джемперли |
| Durvalumab | Дурвалумаб | Imfinzi | Имфинзи |
| Edrecolomab | Эдреколомаб | Panorex | Панорекс |
| Elotuzumab | Элотузумаб | Empliciti | Эмплисити |
| Elranatamab | Элранатамаб | Elrexfio | Элрексфио |
| Emapalumab (emapalumab-lzsg) | Эмапалумаб | Gamifant | Гамифант |
| Enfortumab vedotin (enfortumab vedotin-ejfv) | Энфортумаб ведотин | Padcev | Падцев Онко |
| Enlonstobart | Энлонстобарт | Enshuxing | |
| Envafolimab | Энвафолимаб | Enweida | |
| Epcoritamab | Эпкоритамаб | Tepkinly, Epkinly | Тепкинли, Эпкинли |
| Gemtuzumab ozogamicin | Гемтузумаб озогамицин | Mylotarg | Милотарг |
| Glofitamab | Глофитамаб | Columvi | Колумви |
| Ibritumomab tiuxetan | ибритумомаб тиуксетан | Zevalin | Зевалин |
| Inotuzumab ozogamicin | Инотузумаб озогамицин | Besponsa | Биспонса |
| Ipilimumab | Ипилимумаб | Yervoy | Ервой |
| Isatuximab (isatuximab-irfc) | Изатуксимаб | Sarclisa | Сарклиза |
| Loncastuximab tesirine | Лонкастуксимаб тесирин | Zynlonta | Зинлонта |
| Margetuximab-cmkb | Маргетуксимаб | Margenza | Мардженза, Маргенза |
| Mirvetuximab soravtansine | Мирветуксимаб соравтанзин | Elahere | Элахер (Элахере) |
| Mogamulizumab (mogamulizumab-kpkc) | Могамулизумаб | Poteligeo | Потелигео, Потелиджео |
| Mosunetuzumab | Мосунетузумаб | Lunsumio | Лансумио |
| Moxetumomab pasudotox (moxetumomab pasudotox-tdfk) | Моксетумумаб пасудотокс | Lumoxiti | Люмоксити |
| Naxitamab-gqgk | Накситамаб-гкгк | Danyelza | Даниэлза |
| Necitumumab | Нецитумумаб | Portrazza | Портразза, Портрацца |
| Nimotuzumab | Нимотузумаб | TheraCIM, Biomab-Egfr | |
| Nivolumab | Ниволумаб | Opdivo | Опдиво |
| Inetetamab (cipterbin) | Инететамаб | Cipterbin | |
| Obinutuzumab | Обинутузумаб | Gazyva, Gazyvaro | Газива, Газиваро |
| Ofatumumab | Офатумумаб | Arzerra | Арзерра |
| Olaratumab | Оларатумаб | Lartruvo | Лартруво |
| Panitumumab | Панитумумаб | Vectibix | Вектибикс |
| Pembrolizumab | Пембролизумаб | Keytruda | Китруда, Арфлейда, Пемброриа, Пембролизумаб |
| Penpulimab | Пенпулимаб | Anniko | |
| Pertuzumab | Пертузумаб | Perjeta | Перьета |
| Polatuzumab vedotin (polatuzumab vedotin-piiq) | Полатузумаб ведотин | Polivy | Полайви |
| Prolgolimab | Пролголимаб | Forteca | Фортека |
| Pucotenlimab | Пукотенлимаб | Puyouheng | – |
| Racotumomab | Ракотумомаб | Vaxira | Ваксира |
| Ramucirumab | Рамуцирумаб | Cyramza | Цирамза |
| Relatlimab | Релатлимаб | Opdualag (relatlimab + nivolumab combo) | Опдуалаг |
| Retifanlimab | Ретифанлимаб | Zynyz | Зиниз, Зыниз |
| Ripertamab | Anpingxi | ||
| Rituximab | Ритуксимаб | MabThera, Rituxan | Мабтера, Ритуксан, Ацеллбия, Реддитукс, Ритуксара, Ритуксимаб |
| Sacituzumab govitecan (sacituzumab govitecan-hziy) | Сацитузумаб говитекан | Trodelvy | Тродельви |
| Serplulimab | Серплюлимаб | Hetronifly, Hansizhuang | |
| Sintilimab | Синтилимаб | Tyvyt | Тивит |
| Socazolimab | Соказолимаб | Shan Keyu | |
| Sugemalimab | Сугемалимаб | Cejemly | |
| Tafasitamab (tafasitamab-cxix) | Тафаситамаб | Monjuvi, Minjuvi | Монджуви |
| Tagitanlimab | Тагитанлимаб | ||
| Talquetamab (talquetamab-tgvs) | Талкетамаб | Talvey | Талви |
| Tebentafusp | Тебентафусп | Kimmtrak | Киммтрак |
| Teclistamab | Теклистамаб | Tecvayli | Теквэйли |
| Tislelizumab | Тислелизумаб | Tevimbra | Тевимбра |
| Tisotumab vedotin, tisotumab vedotin-tftv | Тисотумаб Ведотин-тфтв | Tivdak | Тивдак |
| Toripalimab | Торипалимаб | Loqtorzi, Tuoyi | Локторзи |
| Tositumomab-I131 | Тоситумомаб | Bexxar | Бексар |
| Trastuzumab | Трастузумаб | Herceptin | Гертикад, Герцептин, Тразимера, Трастузумаб |
| Trastuzumab deruxtecan | Трастузумаб дерукстекан | Enhertu | Энхерту |
| Trastuzumab duocarmazine | трастузумаб дуокармазин | ||
| Trastuzumab emtansine | Трастузумаб эмтанзин | Kadcyla | Кадсила |
| Tremelimumab | Тремелимумаб | Imjudo | Имджудо |
| Zimberelimab | Зимберелимаб | YuTuo | |
| Zolbetuximab | Золбетуксимаб | Vyloy | |
| Zuberitamab | Зуберитамаб | Enrexib | |
| Odronextamab | Одронекстамаб | Ordspono | |
| Ivonescimab | Ивонесцимаб | ||
| Benmelstobart | Бенмелстобарт | ||
| Trastuzumab botidotin | Трастузумаб ботидотин | ||
| Iparomlimab | Ипаромлимаб | ||
| Iparomlimab, tuvonralimab | Ипаромлимаб, Тувонралимаб | ||
| Tarlatamab | тарлатамаб | Imdelltra | Имделлтра |
| Patritumab deruxtecan | Патритумаб дерукстекан | ||
| Sacituzumab tirumotecan | Сацитузумаб тирумотекан | ||
| Zanidatamab | Занидатамаб | ||
| Linvoseltamab | Линвосельтамаб | ||
| Datopotamab deruxtecan | Датопотамаб дерукстекан | ||
| Zenocutuzumab | Зенокутузумаб | ||
| Nurulimab | Нурулимаб | Nurdati | Нурдати |
| Suvemcitug | Сувемцитуг | ||
| Bifikafusp alfa, Onfekafusp alfa | Бификафусп Альфа Онфекафусп альфа | ||
| Telisotuzumab vedotin | Телисотузумаб Ведотин |
Для лечения аутоиммуных и других болезней, связанных с иммунитетом, используют следующие МАТ (кликните, чтобы увидеть таблицу):
| INN | МНН | Brand Name | Наименование препарата |
| Adalimumab | Адалимумаб | Humira | Хумира, Адалимумаб, Далибра, Эксэмптия |
| Anifrolumab, anifrolumab-fnia | Анифролумаб | Saphnelo | Сафнело |
| Basiliximab | Базиликсимаб | Simulect | Симулект |
| Batoclimab | Батоклимаб | ||
| Belimumab | Белимумаб | Benlysta | Бенлиста |
| Benralizumab | Бенрализумаб | Fasenra | Фазенра |
| Bimekizumab (bimekizumab-bkzx) | Бимекизумаб-бкзкс | Bimzelx | Бимзелекс,Бимцелкс |
| Brodalumab | Бродалумаб | Siliq, Lumicef, Kyntheum | Силик |
| Caplacizumab (caplacizumab-yhdp) | Каплацизумаб | Cablivi | Кабливи |
| Certolizumab pegol | Цертолизимаб пэгол | Cimzia | Симзия |
| Daclizumab | Даклизумаб | Zinbryta; Zenapax | Зинбрита Зенапакс |
| Divozilimab | Дивозилимаб | Ivlizi | Ивлизи |
| Dupilumab | Дупилумаб | Dupixent | Дупиксент |
| Efalizumab | Эфализумаб | Raptiva | Раптива |
| Golimumab | Голимумаб | Simponi | Симпони |
| Guselkumab | Гуселькумаб | Tremfya | Тремфея |
| Inebilizumab (inebilizumab-cdon) | Инебилизумаб | Uplizna | Уплизна |
| Infliximab | Инфликсимаб | Remicade | Ремикейд |
| Itolizumab | Итолизумаб | Alzumab | |
| Ixekizumab | Иксекизумаб | Taltz | Талс |
| Lebrikizumab | Лебрикизумаб | Ebglyss | Эбглисс |
| Levilimab | Левилимаб | Ilsira | Илсира |
| Mepolizumab | Меполизумаб | Nucala | Нукала |
| Mirikizumab | Мирикизумаб | Omvoh | Омвох |
| Muromonab-CD3 | Муромонаб CD3 | Orthoclone Okt3 | Ортоклон ОКТ-3 |
| Natalizumab | Натализумаб | Tysabri | Тизабри |
| Nemolizumab (nemolizumab-ilto) | Немолизумаб | Mitchga, Nemluvio | Немлувио |
| Netakimab | Нетакимаб | Efleira | Эфлейра |
| Ocrelizumab | Окрелизумаб | Ocrevus | Окревус |
| Olokizumab | Олокизумаб | Artlegia | Артлегиа |
| Omalizumab | Омализумаб | Xolair | Ксолар, Генолар, Омализумаб |
| Ozoralizumab | Озорализумаб | Nanozora | Нанозора |
| Reslizumab | Реслизумаб | Cinqaero, Cinqair | Синкейро |
| Risankizumab (risankizumab-rzaa) | Рисанкизумаб | Skyrizi | Скайризи |
| Rozanolixizumab | Розаноликсизумаб | Rystiggo | Ристигго |
| Sarilumab | Сарилумаб | Kevzara | Кевзара |
| Satralizumab (satralizumab-mwge) | Сатрализумаб | Enspryng | Энспринг |
| Secukinumab | Секукинумаб | Cosentyx | Козэнтикс |
| Siltuximab | Силтуксимаб | Sylvant | Силвант |
| Spesolimab | Спесолимаб | Spevigo | Спевиго |
| Sutimlimab (sutimlimab-jome) | Сутимлимаб | Enjaymo | Энджаймо |
| Teplizumab | Теплизумаб | Tzield | Тизилд |
| Tezepelumab | Тезепелумаб | Tezspire | Тезспире |
| Tildrakizumab (tildrakizumab-asmn) | Тилдракизумаб | Ilumya | Илюмиа |
| Tocilizumab | Тоцилизумаб | RoActemra, Actemra | Актемра, Компларейт |
| Tralokinumab | Тралокинумаб | Adtralza | Адтралза |
| Ublituximab | Ублитуксимаб | Briumvi | Бриумви |
| Ustekinumab | Устекинумаб | Stelara | Стелара,Стилейкин |
| Vedolizumab | Ведолизумаб | Entyvio | Энтивио |
| Vunakizumab | Вунакизумаб | ||
| Ebdarokimab | Эбдарокимаб | ||
| Suciraslimab | Суцираслимаб | ||
| Xeligekimab | Кселигекимаб | ||
| Stapokibart | Стапокибарт | Kangyueda | |
| Axatilimab, axatilimab-csfr | Аксатилимаб | Niktimvo | Никтимво |
| Seniprutug | Сенипрутуг | Tribuvia | Трибувиа |
| No INN; Genakumab | Генакумаб | ||
| Nipocalimab | Нипокалимаб | ||
| Picankibart | Пиканкибарт |
Для лечения генетических, инфекционных, кардиологических, неврологических, офтальмологических заболеваний разработаны препараты на основе следующих антител (кликните, чтобы увидеть таблицу):
| INN | МНН | Brand Name | Наименование препарата |
| Abciximab | Абциксимаб | Reopro | РеоПро |
| Aducanumab | Адуканумаб | Aduhelm | Адухельм |
| Alirocumab | Алирокумаб | Praluent | Пралуэнт |
| Amubarvimab + Romlusevimab | Амубарвимаб + Ромлусевимаб | ||
| Ansuvimab-zykl | Ансувимаб | Ebanga | Эбанга |
| Atoltivimab, maftivimab, and odesivimab-ebgn | Атолтивимаб, мафтивимаб, одесивимаб | Inmazeb | Инмазеб |
| Bezlotoxumab | Безлотоксумаб | Zinplava | Зинплава |
| Brolucizumab (brolucizumab-dbll) | Бролуцизумаб | Beovu | Беовью Биову, Беову, Визкью |
| Burosumab (burosumab-twza) | Буросумаб | Crysvita | Крисвита |
| Canakinumab | Канакинумаб | Ilaris | Иларис |
| Casirivimab + imdevimab | Касиривимаб + Имдевимаб | Regen-COV, Ronapreve | Ронапрев Ронаприв |
| Concizumab | Концизумаб | Alhemo™ | Алхемо |
| Crizanlizumab (crizanlizumab-tmca) | Кризанлизумаб | Adakveo | Адаквио Адаквео |
| Crovalimab | Кровалимаб | PiaSky | ПайяСкай ПиаСкай |
| Denosumab | Деносумаб | Prolia Xgeva | Пролиа Эксджива |
| Docaravimab and Miromavimab | Докаравимаб + Миромавимаб | Twinrab | Твинраб |
| Donanemab | Донанемаб | Kisunla | Кисунла |
| Ebronucimab | Эбронуцимаб | ||
| Eculizumab | Экулизумаб | Soliris | Солирис |
| Emicizumab | Эмицизумаб | Hemlibra | Гемлибра |
| Eptinezumab (eptinezumab-jjmr) | Эптинезумаб | Vyepti | Вайэпти |
| Erenumab (erenumab-aooe) | Эренумаб | Aimovig | Аймовиг Иринэкс |
| Evinacumab | Эвинакумаб | Evkeeza | Эвкиза |
| Evolocumab | Эволокумаб | Repatha | Репата |
| Faricimab, faricimab-svoa | Фарицимаб | Vabysmo | Вабисмо |
| Fremanezumab (fremanezumab-vfrm) | Фреманезумаб | Ajovy | Аджови |
| Galcanezumab (galcanezumab-gnlm) | Галканезумаб | Emgality | Эмгалити |
| Ibalizumab (ibalizumab-uiyk) | Ибализумаб | Trogarzo | Трогарзо |
| Idarucizumab | Идаруцизумаб | Praxbind | Праксбайнд |
| Lanadelumab (lanadelumab-flyo) | Ланаделумаб | Takhzyro | Такзайро |
| Lecanemab | Леканемаб | Leqembi | Лекемби |
| Narlumosbart | Нарлумосбарт | Jinlitai | |
| Narsoplimab | Нарсоплимаб | ||
| Nebacumab | Небакумаб | Centoxin | Центоксин |
| Nirsevimab | Нирсевимаб | Beyfortus | Бейфортус |
| No INN; RabiShield | Рабишилд | RabiShield | |
| Obiltoxaximab | Обилтоксаксимаб | Anthim | |
| Ongericimab | Онгерицимаб | ||
| Ormutivimab | Ормутивимаб | ||
| Pabinafusp alfa | Пабинафусп альфа | Izcargo | |
| Palivizumab | Паливизумаб | Synagis | Синагис |
| Pozelimab | Позелимаб | Veopoz | Виопоз Веопоз |
| Ranibizumab | Ранибизумаб | Lucentis | Луцентис Лаксолан Рамилатис |
| Ravulizumab (ravulizumab-cwvz) | Равулизумаб | Ultomiris | Ултомирис |
| Raxibacumab | Раксибакумаб | ||
| Regdanvimab | Регданвимаб | Regkirona | Регкирона |
| Romosozumab (romosozumab-aqqg) | Ромосозумаб | Evenity | Ивенити |
| Sotrovimab | Сотровимаб | Xevudy | Ксевуди |
| Tafolecimab | Тафолецимаб | Sintbilo | Синтбило |
| Teprotumumab (teprotumumab-trbw) | Тепротумумаб | Tepezza | Тепезза |
| Tixagevimab, cilgavimab | Тиксагевимаб + Цилгавимаб | Evusheld | Эвушелд |
| Recaticimab | Рекатицимаб | ||
| Vilobelimab | Вилобелимаб | Gohibic | Гохибик |
| Marstacimab | Марстацимаб | Hympavzi | Химпавзи |
| Garadacimab | Гарадацимаб | ||
| TNM002 | |||
| Bentracimab | Бентрацимаб | ||
| Mazorelvimab, Zamerovimab | Мазорелвимаб Замеровимаб |